
Rethinking Appetite
Gut–brain pathways that shape appetite: vagal sensory circuits, nutrient identity, and the neural architecture of eating patterns

For most of modern medicine, appetite has been treated as a relatively simple physiological phenomenon. We eat when we are hungry. We stop when we are full. If weight gain occurs, it is because caloric intake exceeds energy expenditure.
This framework has been extraordinarily productive. It led to the discovery of hormones such as leptin, ghrelin, CCK, and GLP-1, and to the identification of hypothalamic circuits that regulate energy balance. It also shaped how obesity has been conceptualized for decades, as a disorder of dysregulated intake relative to metabolic demand.
Yet this model largely focuses on hormones and brain centers that regulate energy balance. It says less about how the gut senses a meal and conveys information to the brain. Emerging evidence suggests that the vagus nerve is not simply a conduit for fullness signals, but a sensory interface that carries multiple dimensions of feeding including volume, nutrient identity, and internal state. The gut routes different aspects of a meal through specialized neural circuits that influence both physiology and cognition, including motivation, reinforcement, and memory.
Mapping the Sensory Interface
How did we learn that the vagus carries different kinds of information from the gut?
The answer emerged gradually, through a sequence of anatomical, molecular and functional studies. The gastrointestinal tract is densely innervated by vagal sensory terminals whose cell bodies sit just outside the brain. These neurons extend specialized endings into the stomach and small intestine, where they detect both mechanical stretch and chemical signals generated during digestion. Long before molecular tools were available, anatomical studies by Powley, Berthoud, and others identified distinct vagal terminal structures, including intraganglionic laminar endings (IGLEs) embedded within the muscle layers of the gut wall and mucosal endings that innervate the intestinal villi. These morphologically distinct structures suggested that vagal neurons might be specialized to detect different features of a meal.
A major advance came in 2016, when the Liberles lab demonstrated this anatomical diversity corresponded to genetically distinct sensory neurons. Neurons expressing GLP1R preferentially formed IGLE terminals consistent with stretch detection, whereas neurons expressing GPR65 innervated the intestinal mucosa, consistent with chemosensory roles. This work linked molecular identity to anatomy and established that the vagus nerve transmits at least two fundamentally different types of information about how much has been eaten and what has been eaten.
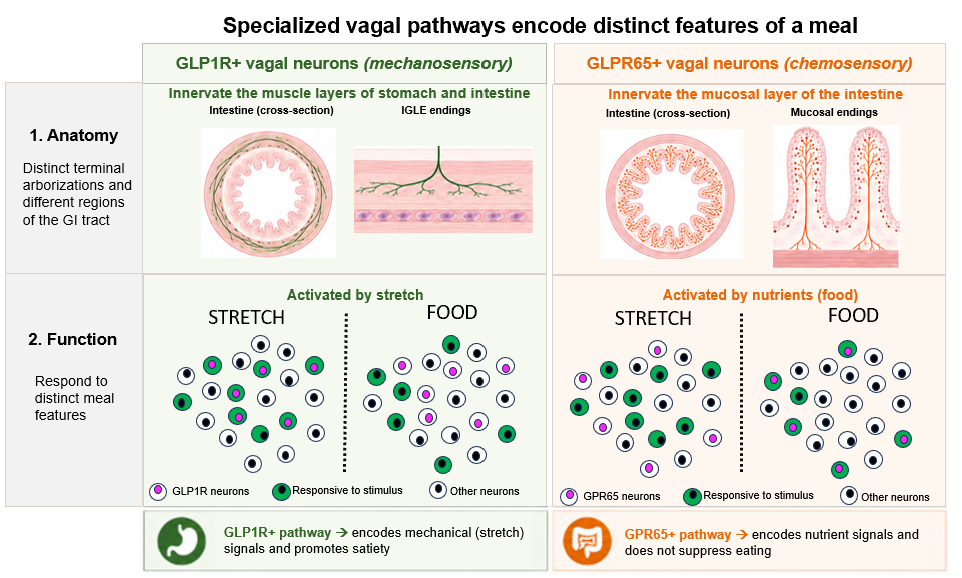
Single-cell RNA sequencing soon refined this picture further. The Knight laboratory identified additional vagal sensory clusters and generated mouse lines that allowed their terminals and functions to be mapped directly. Within the mechanosensory class, two IGLE-forming populations were identified: the previously described GLP1R-expressing neurons and a newly characterized Oxtr-expressing population. Optogenetic stimulation of either population reduced food intake and promoted meal termination, providing causal evidence that stretch-sensitive vagal pathways signal fullness to the brain. In contrast, stimulation of chemosensory populations did not acutely suppress feeding, suggesting that these neurons participate in different aspects of eating behavior.
A Gut-Reward Circuit
If these chemosensory neurons do not terminate a meal, how do they contribute to eating behavior?
In 2018, we activated right-sided vagal sensory neurons innervating the upper gastrointestinal tract. This engaged a defined pathway from the intestine through two brainstem hubs to dopamine-producing neurons in the midbrain. Using optogenetic stimulation to mimic the intestinal input to the brainstem hubs, we observed increased dopamine neuron activity and behavioral reinforcement.
Importantly, these effects were independent of taste. Nutrients delivered directly into the intestine, bypassing oral sensory input, were sufficient to drive reinforcement. The gut itself could signal reward!
Animals learned to self-stimulate this gut-vagal pathway. In real-time place preference experiments, activation of the circuit caused animals to spend more time in the stimulation-paired chamber. In flavor-nutrient conditioning paradigms, pairing circuit activation with a neutral flavor was sufficient to induce learned preference, whereas, inhibiting the pathway during nutrient infusion prevented reinforcement learning. Importantly, these effects were independent of taste. Nutrients delivered directly into the intestine, bypassing oral sensory input, were sufficient to drive reinforcement. The gut itself could signal reward!
Parallel Nutrient Pathways and Motivational Drive
Since gut circuitry engages brain reward regions, we wondered which specific intestinal signals might provide motivation to eat. Could fats and sugars, the defining components of many obesogenic foods, recruit this brain reward pathway?
Using in vivo calcium imaging during controlled nutrient infusion into the gut, we were surprised to find that fats and sugars do not converge onto a single chemosensory population. Instead, they activate distinct subsets of vagal sensory neurons. These fat-responsive and sugar-responsive populations are anatomically separable and project through parallel pathways that drive dopamine release in the midbrain
To determine whether these populations are functionally distinct, we used a mouse genetic tool for activity-dependent labeling (FosTRAP mice) to selectively target neurons activated by fat or sugar. Deletion of fat-responsive neurons abolished reinforcement learning for flavors paired with intestinal fat infusion but did not impair sugar-driven learning. Conversely, deletion of sugar-responsive neurons disrupted sugar reinforcement while leaving fat reinforcement intact. These results indicate that fat and sugar are encoded by separate vagal circuits, each capable of independently driving reinforcement learning for its associated nutrient..
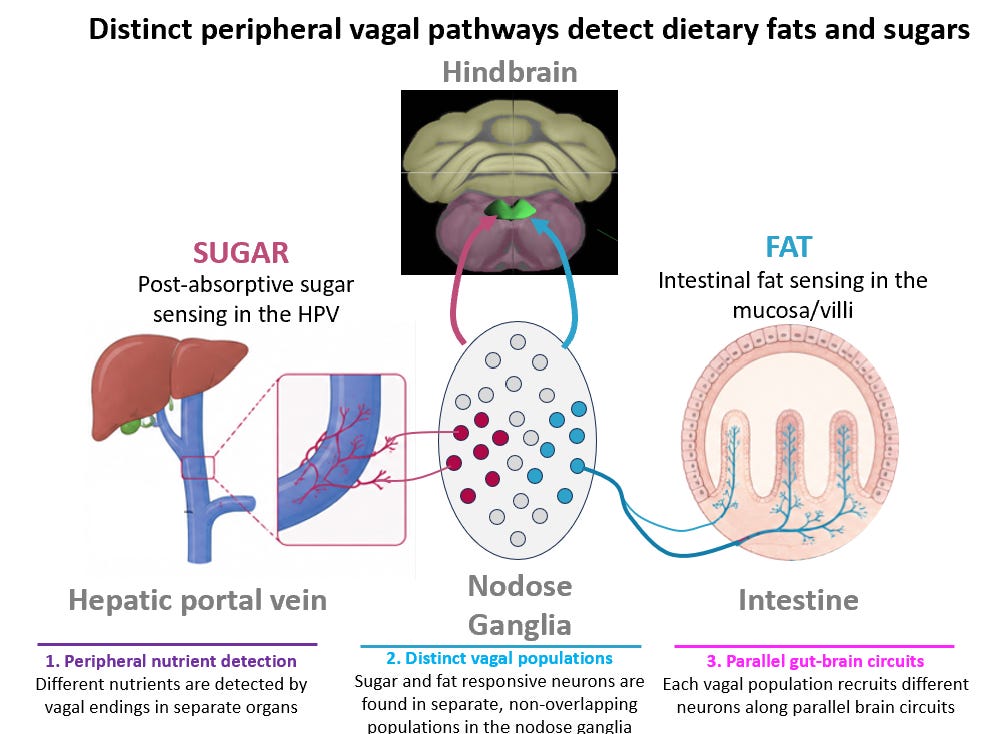
We next asked whether activating these circuits is sufficient to promote motivation. Optogenetic stimulation of either fat-responsive or sugar-responsive pathways was sufficient to support self-stimulation, demonstrating that activation of each circuit is intrinsically reinforcing. In other words, the gut contains parallel channels that can independently generate motivational drive for specific macronutrients.
If separate circuits independently promote reinforcement, we reasoned that their combined activation might amplify motivational drive. To test this, we gave mice access to solutions containing fat alone, sugar alone, or a combination of fat and sugar, carefully matched for total caloric content. Despite equivalent calories, mice consumed nearly twice as much of the combined solution compared to either nutrient alone. Consistent with this behavioral effect, the fat-sugar combination produced greater dopamine release than a single macronutrient. When fat and sugar are consumed together, the gut engages multiple reinforcement circuits at once, producing a stronger motivational signal than either nutrient alone.
In nature, fat-rich and sugar-rich foods are typically distinct. Thus, separate reinforcement systems may have evolved to ensure motivation to seek out scarce energy sources. However, in modern food environments, processed foods frequently combine both macronutrients within the same food.
In nature, fat-rich and sugar-rich foods are typically distinct. Thus, separate reinforcement systems may have evolved to ensure motivation to seek out scarce energy sources. However, in modern food environments, processed foods frequently combine both macronutrients within the same food. Simultaneous recruitment of parallel gut-reward circuits may therefore increase motivational drive and vulnerability to overconsumption.
Importantly, similar findings have been reported in humans. Dana Small’s lab showed that foods combining fat and sugar elicit greater activity in reward-related brain regions than foods containing either macronutrient alone, and participants are willing to pay more for these combined foods. This convergence across species suggests that parallel nutrient reward circuits are evolutionarily conserved.
From Motivation to Memory
How do these reinforcement signals shape food choice over time?
Reinforcement increases the motivational value of a food in the moment. For those effects to shape future intake, however, they must be encoded and stored. Eating behavior is shaped not only by current physiological need, but also by remembered outcomes of prior meals.
The hippocampus has long been recognized for its role in contextual and spatial learning, and prior studies have suggested that it influences food seeking behavior. Until recently, however, its direct role in controlling food intake remained unclear.
We asked whether gut-derived nutrient signals engage hippocampal circuits to influence spatial memory and eating. Using activity-dependent labeling and circuit-specific manipulations, we found that nutrient-responsive vagal pathways activate defined hippocampal ensembles. Deleting these hippocampal neurons impaired spatial memory for nutrient location and selectively reduced intake of that specific nutrient without affecting consumption of others. Conversely, stimulation of the same circuit enhanced memory recall and increased intake of the associated nutrient.
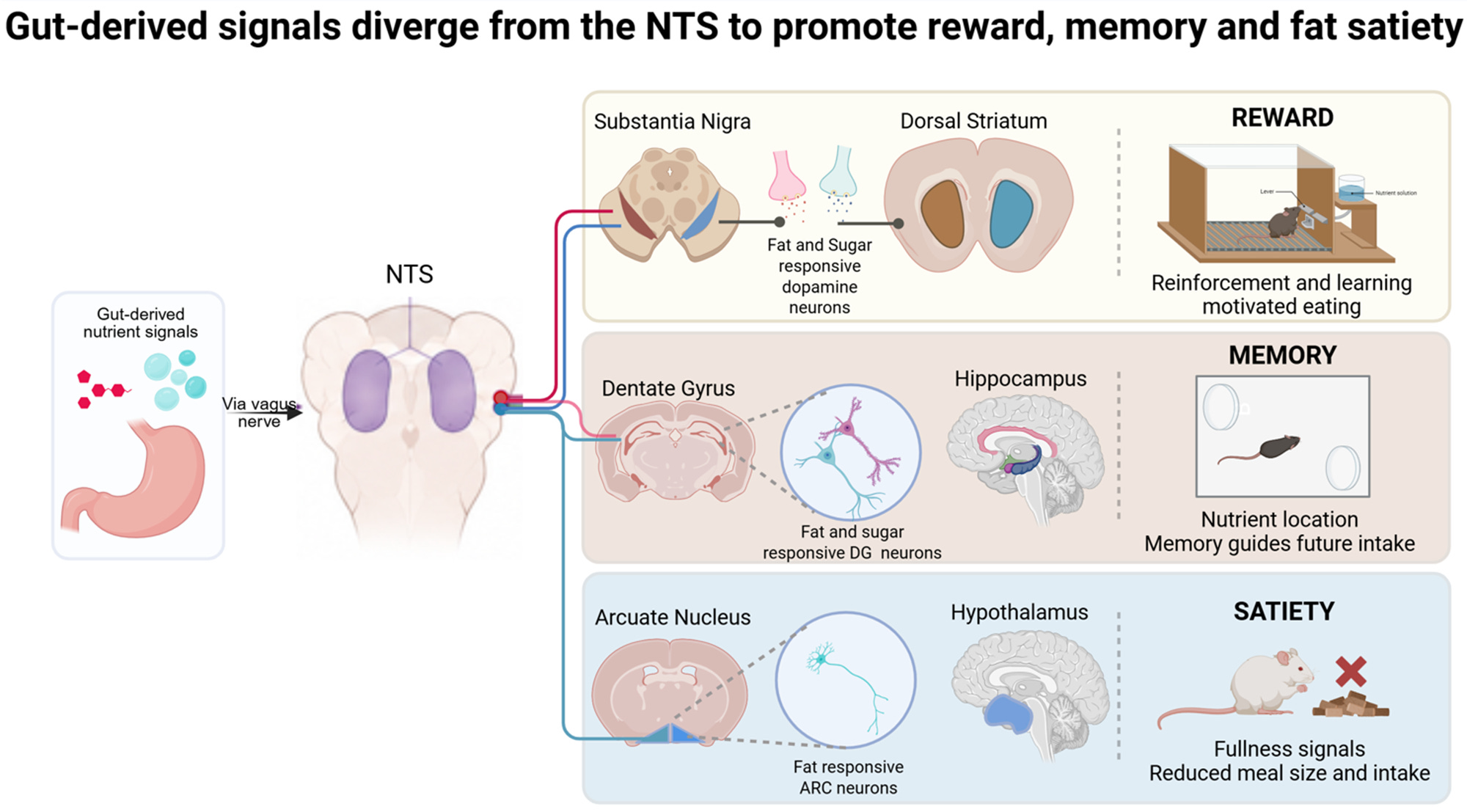
These experiments demonstrate that hippocampal circuits are not merely involved in seeking food but are necessary for regulating nutrient-specific intake. Importantly, disruption of this pathway altered feeding behavior in models of obesity, indicating that gut-to-hippocampus signaling contributes to the control of body weight. These findings extend the function of the gut-brain axis beyond satiety and reinforcement to the spatial and contextual consequences of eating, thus shaping future consumption patterns in a nutrient-specific manner.
When the System Breaks Down
What happens when the flow of information from the gut to the brain is disrupted?
Obesity is often described as a disorder of excess intake relative to expenditure. Within the framework of a layered gut-brain architecture, however, it may also reflect impaired sensory communication. If stretch-sensitive neurons fail to accurately convey fullness, meal termination may be delayed. If nutrient-responsive pathways are altered, reinforcement learning may no longer reflect true metabolic benefit. If gut-derived signals fail to properly engage memory circuits, the ability to use prior physiological experience to guide intake may be compromised. In addition, if these signals are normally partitioned across parallel pathways, disruption may not simply increase or decrease appetite uniformly. It may alter the balance between physiological regulation and motivational drive, or between internal state and remembered outcome.
Future strategies may therefore require not only suppressing hunger signals, but also restoring accurate sensory communication between the gut and brain.
The remarkable success of GLP-1 receptor agonists underscores the therapeutic power of targeting gut-brain pathways. These drugs effectively reduce appetite and promote weight loss by engaging central satiety circuits while also reducing the motivational pull of food. In doing so, they may compensate for impaired peripheral signaling and restore control over food intake. However, weight regain after discontinuation indicates that while GLP-1 receptor agonists effectively dampen appetite, they do not fundamentally repair the neural circuits that integrate satiety, reinforcement, and memory. Future strategies may therefore require not only suppressing hunger signals, but also restoring accurate sensory communication between the gut and brain.
Vagal Afferent Signaling in Eating and Beyond
By understanding obesity as a disorder of sensory integration, rather than solely of willpower or caloric excess, we shift attention toward the neural circuits that interpret information from the intestine. Restoring or recalibrating these pathways may complement pharmacological advances and improve long-term outcomes.
By understanding obesity as a disorder of sensory integration, rather than solely of willpower or caloric excess, we shift attention toward the neural circuits that interpret information from the intestine.
Much of my scientific work has focused on how the vagus nerve carries information about the body’s internal state to the brain. What has become increasingly clear is that these signals do more than regulate hunger or fullness. They shape how the brain assigns value to food, how it stores memories of what we have eaten, and how internal state influences future behavior. This broader view of vagal signaling may have implications not only for obesity and metabolic disease, but also for conditions in which motivation, learning, and cognition are altered.
Editor’s Note This month’s article illustrates how frontier neuroscience can map autonomic neural pathways in detail from body to brain, and sheds light on the complex dynamics and dimensions of appetite. While much of this work was motivated by the public health challenge of obesity, it personally made me think of puzzles I’ve faced at times with stubborn underweightness and broad food intolerances. It also illustrates how the ANS interconnects with slightly higher brain regions accessible to the conscious mind, where autonomic transitions into “automatic”, such as the centers for habit and memory. These shape much of our day-to-day lives and, in turn, reshape our autonomic lives and our bodily physiology. The plot thickens. -Vijay Iyer
 | A guest post by
|